Genetic Analysis of Traits Related to Leaf Sheath in Rice (Oryza sativa L.) 
2. Hangzhou Normal University, Hangzhou, 310036, P.R. China
* These authors contributed equally to this paper
 Author
Author  Correspondence author
Correspondence author
Rice Genomics and Genetics, 2011, Vol. 2, No. 3 doi: 10.5376/rgg.2011.02.0003
Received: 23 Nov., 2011 Accepted: 07 Dec., 2011 Published: 31 Dec., 2011
Liu et al., 2011, Genetic Analysis of Traits Related to Leaf Sheath in Rice (Oryza sativa L.), Molecular Plant Breeding, 9(3): 278-287 (doi: 10.3969/mpb.009.000278)
The double haploid (DH) population derived from a typical cross between an indica rice TN1 and a japonica rice CJ06 was used in this research to investigate the genetic characteristics of length, thickness and dry weight of leaf sheath among individuals of the DH population as well as their parents. The results showed that the sheath length in rice was a trait with high heritability and less environmental impacts, as well the sheath length was demonstrated significantly positive correlation with the sheath thickness and the dry sheath weight. There were 31 QTLs related to the trait of rice sheath to be detected, which were mapping on the chromosome 1, 4, 6, 8, 10 and 11, respectively. The LOD values of these QTLs were in range from 2.58 to 4.81, and the phenotypic variations were in range from 7.5% to 35.3%. The interval of RM548-RM6997 located on chromosome 4 in rice genome was considered as a major effective QTL locus, which might confer the traits of rice sheath length, dry weight of sheath and sheath thickness. Comprehensive comparison was conducted between the sheath-related traits and some yield related traits, the results indicated that photosynthetic function of sheath and or synthetic substance transportation of sheath might be closely related to rice yield formation.
Rice leaf sheath is the sheath-like organic part that wraps up the stem node and internodes of the stem around the leaf base, which is derived from lower part of cells of leaf primordia and has functions for protecting intermediate stem meristem and strengthening the mechanical support as well as protecting axillary buds (Lu, 1992; Yao, 2008). Rice leaf sheath is considered as a kind of temporary storage organ to store the excess photosynthetic products in rice plants before the heading stage (Perez et al., 1971). Usually, these stored photosynthetic products will be transported to the grains after heading. The weight of stem and sheath of single rice stem is positively correlated with rice yield, which is an important factor affecting the rice yield (Su et al., 1993). The sheath with its own sheath photosynthesis has great contributions to rice yield, up to 10%~20% (Guo et al., 2007). In addition, as a channel connecting leaves and grains, leaf sheath plays an important physiological function for loading and unloading the photosynthetic products during the grain milking stage (Li, etc., 2008). Therefore, the organic characteristics of stem and sheath are known as an important indicator for evaluating quality of rice population and estimating photosynthetic potential of rice plants after heading (Su et al., 1993; Tao et al., 2002; Zhang et al., 2001).
Leaf sheaths are closely related to supporting ability of rice stem and lodging resistant properties. By comparing the morphological characteristics of the profile of broken stem in different varieties, leaf sheath of the lower internode in lodging varieties is quite thin (Wang et al., 1991). The more stems with tightly wrapped sheaths the more strength of lodging resistant stems in rice, as well the more thick sheath wrapped, the stronger the ability of lodging resistant stems (Wang et al., 1998; Rao et al., 2009). However, with the panicle grain being filled from the late heading to grain filling and ripening, the photosynthetic products stored in stem and sheath are constantly transported to rice ears, easily leading to be more susceptible to lodging in rice ripening stage (Ai, 2006). Lodging not only causes the loss of rice yield, but also reduces grain quality, as well as increases the cost of rice production due to the difficulties of harvesting (Kashiwagi et al., 2008). In view of the important influences of rice leaf sheath on yield and lodging resistance, Some studies were carried out including accumulation of non-structural carbohydrates, the dynamic changes of potassium and silicon content, transportation and coordination of sink-source substance, and so on (Nagata et al., 2002; Zhang et al., 2010; Takahashi et al., 2005). However, the genetic basis of traits related to leaf sheath is few to be reported.
This study systematically analyzed the genetic mechanism of the length, dry weight and thick of leaf sheath during the heading stage, which might provide assisted selection markers and theoretical basis directly using for developing and improving super-high-yield and lodging resistant varieties in rice.
1Results and Analysis
1.1 Performance of sheath traits in DH population
According to the phenotypic performance of the length, dry weight and thickness of first, second and third leaf sheath from the panicle top in DH population and their parents (Table 1), there were quite differences among the mentioned traits in the parents. The sheath length of japonica parent CJ06 was longer than that of the indica parent TN1, and also leaf sheath of the CJ06 was looked more heavy and thickness.
In these measured traits, the variation coefficient of sheath length was about 15% far less than that of sheath dry weight of 35.05%~48.62% and that of sheath thickness of 24.25%~34.11%, which indicated that the trait of leaf sheath should has a high heritability less subject to environmental impact. In the DH population, the measured traits of the length, dry weight and thickness of first, second and third leaf sheath from the panicle top exhibited continuous distributions that would be suitable for QTL mapping analysis.
 Table 1 The Phenotypic performance of DH population and their parental lines |
1.2 Correlation analysis of traits
Correlation analysis showed (Table 2) that the sheath related traits represented highly significant positive correlation except for sheath length to sheath thickness and dry weight of the 3rd leaf sheath from the top, the correlation coefficients of these traits varied in range of 0.376 to 0.963. In terms of the investigated traits such as the length, thickness and dry weight of leaf sheath, the correlations of the traits between the 2nd leaf and 3rd leaf from the top had significantly higher than that of other leaves, for example, the correlation coefficient of the sheath length between the 2nd leaf and 3rd leaf from the top was 0.8127, whereas the correlation coefficients were 0.5792 and 0.3952 for the traits between the 1st leaf and 2nd leaf and between 1st leaf and 3rd leaf, respectively. Similarly, the correlations of the traits of dry weight and thickness of leaf sheath between the 2nd leaf and 3rd leaf from the top had also significantly higher than that of other leaves, which might be related to the evidences that the early vegetable growth in rice transformed the late reproductive growth during the growth and development stage of flag leaf in rice. The trait of 2nd leaf sheath also showed high correlation with other traits, such as the length of leaf sheath of 2nd leaf from the top had higher correlations with the dry weight sheath of other leaves than that of 1st and 3rd leaf from the top had, which might explained that the leaf sheath of the 2nd leaf from the top might take some power from the leaf sheath of the 3rd leaf from the top and also provide the main source of nutrition to flag leaf and its leaf sheath.
 Table 2 Correlation coefficients between leaf sheath and plant height |
1.3 QTL analysis
Thirty one QTLs related traits of leaf sheath were detected by using the DH population (Table 3), which distributed on chromosome 1, 4, 6, 8, 10 and 11 (Figure 1). There were 4 QTLs related to sheath length for flag leaf, 3 QTLs for 2nd leaf and 3 QTLs for 3rd leaf, respectively, which mapped on chromosome 1, 4, 6, 8 and 11 with the LOD score between 2.59~4.81 and explained variations between 9.1%~35.3%. The additive effects of the 2nd and 3rd leaf sheath length come from CJ06, whereas the additive effects of the flag leaf sheath length come from TN1 except for CJ06. Of these detected QTLs, QTLs related to the sheath length of three different position leaves were detected on the regions neighboring RM5389 on the long arm of chromosome 1 and RM21-RM3428 interval on chromosome 11, respectively, while only QTLs related to the sheath length of the flag leaf was detected on RM3-RM162 interval on chromosome 6 and RM1376-RM4085 interval on chromosome 8, respectively, the additive effects of the allele come from indica parent TN1. The interval of RM548-RM6997 on Chromosome 4 is a major loci affecting the leaf sheath length of 2nd and 3rd leaf from the top, which might explain their variation 35.3% and 24.3%, respectively.
 Table 3 The QTL analysis of related traits to leaf sheath |
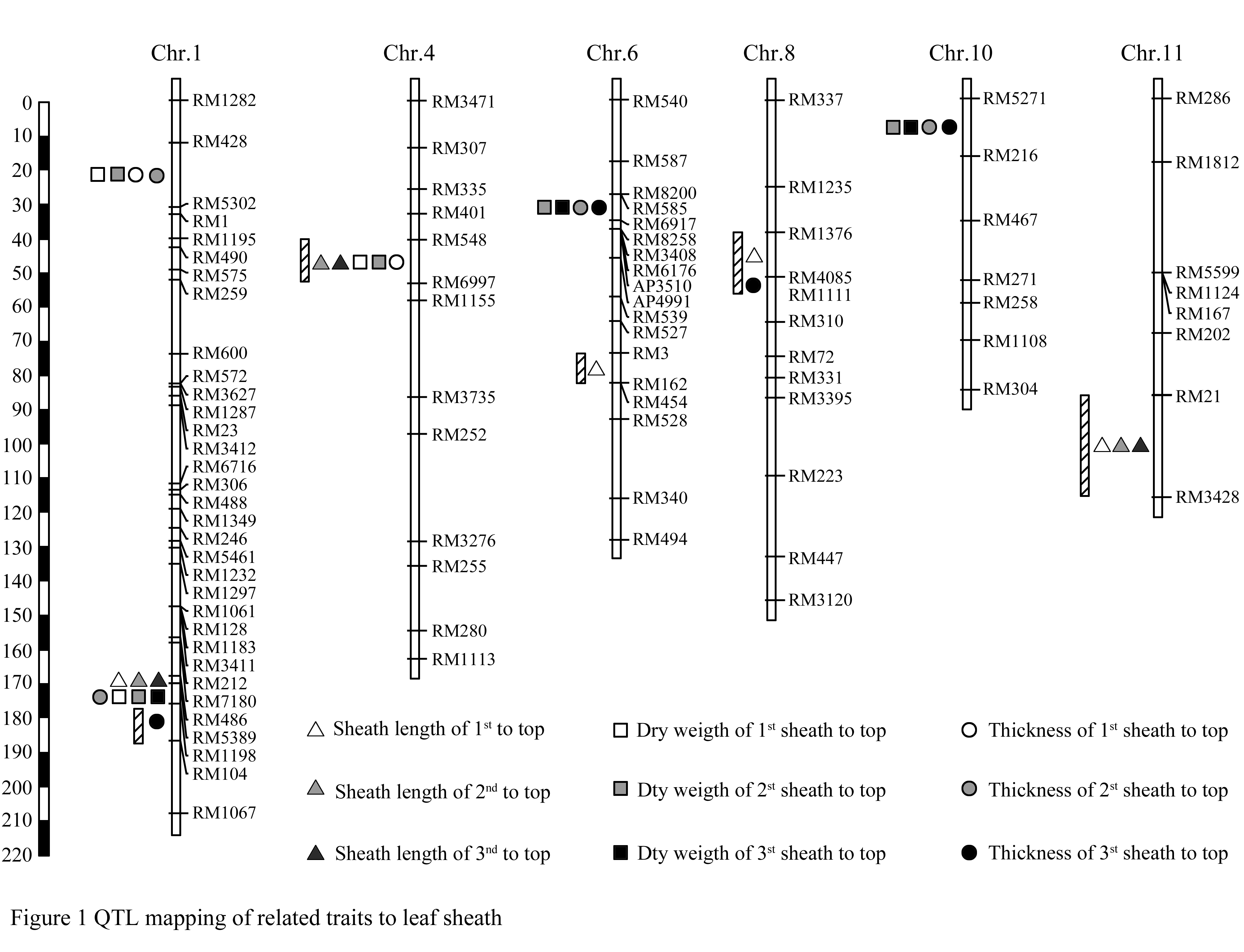 Figure 1 QTL mapping of related traits to leaf sheath |
Eleven QTLs related to dry weigh of sheath were detected in this research, 3 QTLs related to sheath length for flag leaf, 5 QTLs for 2nd leaf and 3 QTLs for 3rd leaf located on chromosome 1, 4, 6 and 10 with LOD values in the range between 2.62~4.12 and explained variation 7.6%~28.6%, respectively. The results indicated that the trait of the dry weight of leaf sheath should be more subjected to environmental influence leading to relative low heritability. As compared these QTLs, QTL locus for leaf sheath dry weight of 2nd leaf sheath covered the QTLs for the sheath dry weight of flag and 3rd leaf, which supported our conclusions that the 2nd leaf from the top played important roles in nutrition storage and transportation. Of these QTLs, QTL locus near RM1198 in the long arm of chromosome 1 had effects on the leaf sheath dry weight of flag leaf, 2nd and 3rd leaf.
There were 2 QTLs related to sheath thickness for flag leaf, 4 QTLs for 2nd leaf and 4 QTLs for 3rd leaf located on chromosome 1, 4, 6, 8 and 10 with LOD values in the range between 2.58~4.54 and explained variation 7.5%~26.0%, respectively. qTT-8 locus for sheath thickness of 3rd leaf from the top and qLT-8 locus for sheath length the flag leaf both located in the neighboring regions in the long arm on chromosome 8, whereas other nine QTLs related to sheath thickness were all mapped on the same regions as QTLs related to the sheath dry weight. The result indicated that leaf sheath thickness mainly linked to leaf sheath dry weight rather than the length of leaf sheath.
1.4 Comparative genetic analysis for QTLs and yield related traits
We also compared the genetic maps between the QTLs related to leaf sheath with some yield related QTLs. The results showed that the QTLs for sheath we detected in this study overlapped or closed some major QTLs related to yield previously reported (Table 4). For example, the sd1 gene leading to the first green revolutionary were overlapped or closed to the QTL regions for the trait of sheath length, thickness and dry weight; An important gene GIF1affecting the rate of grain filling on chromosome 4 was consistent with the major QTLs conferring the traits of sheath length, thickness and dry weight mapped on the RM548-RM6997 region. Further analysis showed that GIF1 gene was quite differences existing between TN1 and CJ06 (Figure 2), Four exons of six exons in GIF1 gene had nucleotide substitutions happened. Such as A nucleotide in first exon of the TN1 had been substituted as G nucleotide from CJ06, resulting in amino acid coding change from Thr to Ala, while these non-synonymous mutations for amino acid coding changes also existed in the 4th and 5th exons of GIF1. There were also the differences of GIF1 between TN1 and CJ06 in the evidences of a large number of intron substitution, deletion or insertion. We also found that the major QTL intervals for leaf sheath related traits we detected were basically consistent with the major yield-related traits QTLs reported previously. For example, many reported yield-related QTLs had been detected in the region or neighborhood of QTLs related to sheath length detected on chromosome 6 and 8, which showed that the photosynthetic functions of leaf sheath or substance transportation might be closely related to yield formation in rice.
 Table 4 The genetic comparisons among the traits related to yield and leaf sheath |
|
Figure 2 Sequence alignment of GIF1 gene between tested cultivars of CJ06 and TN1 |
2 Discussions
Rice leaf sheath plays important roles in early grain development and grain filling, because the sheath can temporarily store nutrients from leaves and roots in early grain filling stage and can synthesize assimilating substance in the mid grain filling stage (Li et al., 2008). In production, a large leaf area index of plants would be helpful to increase the substance assimilation before the heading as well as to enhance the photosynthetic potential after heading. Once the leaf area index maintains stable after heading, increasing the weight of stem and sheath of single plant would be alternative to lift the rice yield. If both of them increasing, the super yield production would be realized (Su et al., 1993). Leaf sheath can delay the aging of stem and increase the strength of the stem, as well as increase plant lodging resistance by enhancing the mechanical strength of stem and sheath in rice (Kashiwagi et al., 2008).
2.1 Relations between leaf sheath and yield
Photosynthesis of the leaf contribute more than 90% crop yield is one of the typical technical theoretical achievements derived from the famous "first green revolution", however, this theory ignored the functions of other "non-leaf photosynthetic organs” , such as sheaths. Su et al (1993) reported that the dry weight of a single stem and sheath of rice in heading stage had positively correlated to the yield of rice. Deshmukh (2008) etc.detected QTLs controlling the 1000-grain weight on chromosomes 1 and 10 by using DH population derived from the cross CT9993-5-10-1-M and IR6226642-6 in different environments. Similarly, Liu et al (1998) detected QTLs related to grain weight, panicle weight and yield on chromosome 1 and 10 by using DH population derived from the cross of m64 and Azucena. In this study we detected two major QTL s conferring the dry weight of leaf sheath on chromosome 1 and 10, which were close to above mentioned QTLs , suggesting that the control the yield in rice. In addition, GIF1, an yielding gene (Wang et al., 2008), mapped on chromosome 4 in the position of 58.6 cM~58.9 cM, where was consistent with the QTLs for the length, dry weight and thickness of leaf sheath of the 1st, 2nd and 3rd leaf from the panicle. This was not only possible that GIF1 loci affect traits of the sheath, but also possible the QTLs we detected would be GIF1. In addition, we also found 12 reported QTLs for yield genes were consistent with our QTL intervals in this research and three reported QTLs were adjacent these region we detected. Therefore, we speculated the traits of leaf sheath, particular in leaf sheath length and sheath thickness, might have great correlations with the rice yield, which had large contributions to enhance rice yield.
2.2 The relations between sheath length and plant height
The expression of genes depends on the environment, interaction between QTL and environment is an important factor in the expression of quantitative traits (Fan, 2001), gene in different environments may have different expression profiles. Zhang et al (2006) thought that the elongation of the rice leaf sheath and internode had a relationship of synchronization, elongation of leaf sheath indirectly affects plant height. Lu et al (1996) and Li et al (2002) detected the QTLs related to plant height on chromosomes 4, 7 and 8 by using DH population derived from the cross of Zhaiyeqing 8 and Jingxi 17 in different environments repeatedly. Ye et al (2006) also detected the QTLs related to plant height located on the chromosome 1 and 4 repeatedly by using F2 and F2:3 populations derived from Pei'ai 64S and Nipponbare in different environments. In this study, we detected two QTLs conferring leaf sheath length on the chromosome 4 in the DH population derived from the cross of CJ 06 and TN1, which overlapped or neighbored the QTL intervals mentioned above, it would have conclusions that the QTLs for plant height were existence of chromosome 4, whether these QTL conferred the traits of leaf sheath length, or these QTLs were located on the same locus, remains to be further validated yet. SD1 is a semi-dwarf gene on the same location in the genome as QTL for sheath thickness of the 3rd leaf from the top, which was consistent with the results of correlation analysis. It implied that theinterval of QTL for sheath thickness might frame probably the sd-1 gene (Lisa et al., 2002).
2.3 The relations between leaf sheath and lodging resistance
Some studies at home and abroad had shown that lodging not only associates with the factors of cropping system , fertilizer and water management, pest and disease control, but also depended on many physical properties, such as plant height, cell wall thickness of stem, dry weight of stem and sheath and so on, besides Hu et al (2008) detected QTLs related to lodging resistance and plant height on chromosome 8 by using the DH population from the cross of Zhaiyeqing 8 and Jingxi 1, whose location was quite close where the QTL for sheath thickness we detected in this experiment. QTL analysis in this study showed that the QTLs for both length and dry weight of leaf sheath in the same interval on the chromosome 4 were detected. Correlation analysis also showed that the traits of leaf sheath length and dry weight had a positive correlation, which predicted that the sheath length might be closely associated with chromosome 4, it needs to further study what kinds of relationship is there and how the association with the traits. Through the comprehensive studies in the genetic maps, there was a QTL for sheath length on chromosome 6 that neighbored the new SCM2 gene released by Ookawa et al (2010) recently, which was the rice gene regulating vascular development and first branch of panicle to affect the yield and rice harvest index and to enhance plant lodging resistance (Terao et al., 2010). We also found that MOC1 was located on the intervals where QTL related to the sheath length of the flag leaf was detected (Li et al., 2003), may fall a long sheath and tiller also has a close relationship. The traits of rice leaf sheath are very complex that closely link to the rice yield, plant height and lodging resistance, it might provide a theoretical reference for high-yield and lodging resistant breeding in future by detecting QTL related to leaf sheath traits.
3 Materials and Methods
3.1 Experiment materials and treatment
A typical japonica cultivar CJ06 mated with a typical indica variety TN1 to generate a hybrid in this research. The anthers of F1 generation were used for in vitro culture, and then went to be diploid by natural or colchicine treatment for homozygous diploid (DH) lines about more than 200 individuals. 120 DH lines with genetic stability were randomly selected to construct molecular linkage map.
3.2 Traits phenotyping
DH population and their parents were sowed on May 10, 2010 in Fuyang experimental station of China National Rice Research Institute, transplanted on June 5, 2010. Each line transplanted 4 rows with six individuals, three repeats were set up in the experiments, field management followed as the general field management in the station. Twenty individuals were collected to be investigated after 20 days behind the heading stage. Three representative plants were selected to be phenotyped the traits of the leaf sheath length (cm) , leaf sheath dry weight (g) and leaf sheath thickness (mg / cm) of 1st leaf (flag leaf) 2nd and 3rd leaf, thereinto, sheath thickness was regarded as the dry weight per unit length of the sheath, calculated as follows:
Sheath thickness (mg / cm) = [leaf sheath dry weight (g) / leaf sheath length (cm)]×1000
At the data were applied to be examined by using statistical software SAS8.0 to figure out the correlations amoung leaf sheath traits.
3.3 Construction of molecular linkage map and QTL mapping
To build the genetic map this DH population, we selected 178 markers from numerous SSRs and STSs that had polymorphism between the parents, which evenly distributed in 12 chromosomes of rice, the genetic constructed by Mapmaker/Exp3.0b in our lab spans1674.8 cM of rice genome, the average distance of the markers is about 9.4 cM, which would be suitable for QTL analysis in general (Zeng et al., 2009; Rao et al., 2010). QTL analysis was applied by mapmaker/QTL1.1B based on LOD score 2.5 to determine whether the marker interval existing, while LOD> 2.5, the highest LOD peak would be indicated the existence of a QTL. The explained variation and additive effect for each QTL was determined, each QTL was assigned a name following the reference by McCouch et al (Khush et al., 1996; McCouch et al., 1997).
3.4 Comparative analysis for QTLs related to traits
According to published reports, we collected some of important genes related to the yield and logging resistance. The first of step was to find the location of these genes in the Gramene QTL Database of on the webpage (http://www.gramene.org/qtl/), and then linked them to our SSR genetic linkage map to compare the locations of genes or QTLs. Finally three cloned genes of sd1, MOC1 and GIF1, and 15 QTLs including GY, GYD (chr.1), nscp, GYD (chr.4), yd6a, yd6, qYl-6-1, GYD (chr.6), gy8, yd8, yld8.1, yld8.2, yld8.3, yld8.4 and tgw11 assigned to the corresponding chromosome.
Authors’ contributions
HJL and YCR are the persons who carried out this experiment; YLY, YJL, LCH, GHZ, JH, participated in some in lab work; LBG, ZYG,LZ, GJD,JL, MXY involved the data analysis and in field work; OQ and DLZ conceived the project and designed the experiments as well as wrote and revised manuscript . All authors had read and agreed the final text.
Acknowledgements
This research is jointly sponsored by the National 973 Project (No. 2007CB10920203), National 863 Project ( No. 30971760) and Excellent youth Team Project in Zhejiang Province to Zeng’s Group (R3090023). Authors thank for two anonymous reviewers with their critical comments. In this paper we mentioned some chemical and reagent suppliers and sequencing service providers, that doesn't mean we would like to recommend or endorse the production of theirs.
References
Ai Z.Y., 2006, Study on relationship between morphological and physiological traits and lodging resistance in super hybrid rice, Thesis for M.S., Agriculture, Hunan University, Supervisor: Ma G.H., pp.13-16
Cho Y.C., Suh J.P., Choi I.S., Hong H.C., Baek M.K., Kang K.H., Kim Y.G., Ahn S.N., Choi H.C., Hwang H.G., and Moon H.P., 2003, QTLs analysis of yield and its related traits in wild rice relative oryza rufipogon, Treat. of Crop Res., 4: 19-29
Deshmukh R., Sonah H., Tiwari A., Mahatman K., Jadhav S., Kadam S., Kesawat M., Reddy N., Deshmukh N., and Verulkar S., 2008, Identification of quantitative trait loci for yield, and related traits in rice, Guangxi Agricultural Sciences, 39(5): 561-564
Fan Y.Y., Zhuang J.Y., Li Q., Sala F., and Zheng K.L., 2001, Analysis of quantitative trait loci (QTL) for plant height and the relation between these QTL and QTL for yield traits in rice, Zuowu Xuebao (Acta Agronomica Sinica), 27(6): 915-922
Guo Y.H., Zhu S.G., Zhang L.B., and Dou H., 2003, Influence on the material characteristics of rice culms in different cultivation conditions, Shenyang Nongye Daxue Xuebao (Journal of Shenyang Agricultural University), 34(1): 4-7
Guo Z.W., Xiao L.T., Luo X.H., Li H.S., Wu C.C., Kang D.L., and Shi Q., 2007, Super hybrid rice "Liangyoupeijiu" photosynthesis flag leaf sheath, Zuowu Xuebao (Acta Agronomica Sinica), 33(9): 1508-1515
Hu J., Fujimoto K., Guo L.B., Zeng D.L., Zhang G.H., Dong G.J., Wang X.H., Zhu L.H., and Qian Q., 2008, QTL analysis of lodging resistance force and lodging resistance-related traits in rice, Zhongguo Shuidao Kexue (Chinese J. Rice Sci.), 22(2): 211-214
Hua J., Xing Y., Wu W., Xu C., Sun X., Yu S., and Zhang Q., 2003, Single-locus heterotic effects and dominance by dominance interactions can adequately explain the genetic basis of heterosis in an elite rice hybrid, Proceedings of the National Academy of Sciences of the United States of America, 100(5): 2574-2579 http://dx.doi.org/10.1073/pnas.0437907100 PMid:12604771 PMCid:151382
Hua J.P., Xing Y.Z., Xu C.G., Sun X.L., Yu S.B., and Zhang Q., 2002, Genetic dissection of an elite rice hybrid revealed that heterozygotes are not always advantageous for performance, Genetics, 162(4): 1885-1895 PMid:12524357 PMCid:1462368
Kashiwagi T., and Ishimaru K., 2004, Identification and functional analysis of a locus for improvement of lodging resistance in rice, Plant Physiol., 134(2): 676-683 http://dx.doi.org/10.1104/pp.103.029355 PMid:14739343 PMCid:344543
Kashiwagi T., Togawa E., Hirotsu N., and Ishimaru K., 2008, Improvement of lodging resistance with QTI S for stem diameter in rice, Theor. Appl. Genet., 117: 749-757 http://dx.doi.org/10.1007/s00122-008-0816-1 PMid:18575836
Khush G.S., 1996, Prospects of and approaches to increasing the genetic yield potential of rice, In: Evenson R.E. (ed.), Rice Research in Asia, Progress and Priorities, CAB International, IRRI, pp.59-71
Lafitte H.R., Courtois B., and Arraudeau M., 2002, Genetic improvement of rice in aerobic systems: Progress from yield to genes, Field-Crops-Research, 75(2-3): 171-190 http://dx.doi.org/10.1016/S0378-4290(02)00025-4
Li R.T., Jiang T.B., Qiu T.Q., Cui C.H., and Gong Z.P., 1996, Study on effect of lodging to yield and relationship between lodging and plant height in rice, Heilongjiang Nongye Kexue (HeiLongjiang Agricultural Science), 1: 13-17
Li S.G., Ma Y.Q., He P., Wang Y.P., Zhou K.D., and Zhu L.H., 2002, Comparative mapping of quantitative trait loci for heading date and plant height in cultivated rice (Oryza sativa L.) across environments, Zuowu Xuebao (Acta Agronomica Sinica), 28(4): 546-550
Li X.H., Xu C.G., Gao Y.J., Yu S.B., Zhang Q., Li J.X., and Tan Y.F., 2000, Analyzing quantitative trait loci for yield using a vegetatively replicated F2 population from a cross between the parents of an elite rice hybrid, Theor. Appl. Genet, 101(1-2): 248-254 http://dx.doi.org/10.1007/s001220051476
Li X.H., Zhang X.W., and Sheng W., 2010, Efforts to promote rice production and promotion of agricultural mechanization and the rapid development of industrialization, Jilin Nongye (Jilin Agricultural), 7: 44-44
Li X.Y., Qian Q., Fu Z.M., Wang Y.H., Xiong G.S., Zeng D.L., Wang X.Q., Liu X.F., Teng S., Hiroshi F., Yuan M., Luok D., Han B., and Li J.Y., 2003, Control of tillering in rice, letters to nature, 10(422): 618-621
Li Z.W., Xiong J., Li Z.F., Qi X.H., Chen H.F., Shao C.H., Wang J.Y., Liang Y.Y., and Lin W.X., 2008, Analysis of diferential expression of proteins in rice leaf sheath during grain filling. Zuowu Xuebao (Acta Agronomica Sinica), 34(4): 619-626 http://dx.doi.org/10.3724/SP.J.1006.2008.00619
Lisa M., Noriyuki K., Rika Y., Junko S., Haruka M., Yumiko M., Masao T., Mizuho S., Shinobu N., and Yuzo M., 2002, Positional cloning of rice semidwarfing gene, sd-1: Rice “Green Revolution Gene” Encodes a Mutant Enzyme Involved in Gibberellin Synthesis, DNA Research, 9(1): 11-17 PMid:11939564
Liu G.F., Lu Y.G., Wang G.C., and Huang N., 1998, Identification of QTL as for plant yield, plant height and their related traits in rice, Huanan Nongye Daxue Xuebao (J. South China Agric Univ.), 19(3): 5-9
Lu C., Shen L., Tan Z., Xu Y., He P., and Chen Y., 1996, Comparative mapping of QTL for agronomic traits of rice across environments using a doubled haploid population, Theor. Appl. Genet., 93(8): 1211-1217 http://dx.doi.org/10.1007/BF00223452
Lu S.W., Xu X.S., and Shen M.J., eds., 1992, Botany, Higher Education Press, Beijing, China, pp.41-43
Ma G.H., Deng O.Y., Wan Y.Z., and Wang X.H., 2000, Resistant physiology to lodging and morphological characters of super hybrid rice I. differences of Si, K and Fiber Contents of the Plant between Pei’ai64S/E32 and Shanyou63, Hunan Nongye Daxue Xuebao (Journal of Hunan Agricultural University), 26(5): 329-331
Ma Y.Y., Wang R.P., Li L., Zuo S.M., Yin Y.J., and Chen Z.X., 2008, Genetics and breeding of rice plant height progress, Henan Nongye Kexue (Henan Agricultural Science), 11: 12-17
Marri P.R., Sarla N., Reddy L.V., and Siddiq E.A., 2005, Identification and mapping of yield and yield related QTLs from an indian accession of oryza rufipogon, BMC Genetics, 6(33): 1-14
PMid:15634360 PMCid:545062
McCouch S.R., Cho Y.G., Yano M., Paul E., and Blinstrnh M., 1997, Repert on QTL nomenclature, Rice Genet. Newel., 14: 11-13
Nagata K., Shimizu H., and Terao T., 2002, Quantitative trait loci for nonstructural carbohydrate accumulation in leaf sheaths and culms of rice and their effects on grain filling, Breeding Science, 52(4): 275-283 http://dx.doi.org/10.1270/jsbbs.52.275
Perez C.M., Pa1miano E.P., Baun L.C., and Juliano B.O., 1971, Starch metabolism in the leaf sheaths and culm of rice, Plant Physiol., 47(3): 404-408 http://dx.doi.org/10.1104/pp.47.3.404
PMid:16657631 PMCid:365878
Rao Y.C., Dong G.J., Zeng D.L., Hu J., Zeng L.J., Gao Z.Y., Zhang G.H., Guo L.B., and Qian Q., 2010, Genetic analysis of leaffolder resistance in rice, J. Genet Genomics, 37(5): 325-31 http://dx.doi.org/10.1016/S1673-8527(09)60050-3
Rao Y.C., Li Y., Dong G.J., Zeng D.L., and Qian Q., 2009, Progress of research on lodging resistance in rice, Zhongguo Daomi (China Rice), 6: 15-19
Takahashi S., Ishimaru K., Yazaki J., Fujii F., Shimbo K., Yamamoto K., Sakata K., Sasaki T., Kishimoto N., and Kikuchi S., 2005, Microarray analysis of sink-source transition in rice leaf sheaths, Breeding Science, 55(2): 153-162 http://dx.doi.org/10.1270/jsbbs.55.153
Tao L.X., and Wang X., 2002, Current status and advances of physiological barrier in sub-species hybrid rice seed setting review of china., Zhongguo Nongye Keji Daobao (Agricultural Science and Technology), 4(1): 54-59
Terao T., Nagata K., Morino K., and Hirose T., 2010, A gene controlling the number of primary rachis branches also controls the vascular bundle formation and hence is responsible to increase the harvest index and grain yield in rice, Theoretical and Applied Genetics, 120(5): 875-893 http://dx.doi.org/10.1007/s00122-009-1218-8 PMid:20151298
Wang E., Wang J.J., Zhu X.D., Hao W., Wang L.Y., Li Q., Zhang L.X., He W., Lu B.R., Lin H.X., Ma H., Zhang G.Q., and He Z.H., 2008, Control of rice grain-filling and yield by a gene with a potential signature of domestication, Nature Genetics, 11(40): 1370-1374 http://dx.doi.org/10.1038/ng.220 PMid:18820698
Wang S.B., 1991, Studies on lodging inrice plant, II-Morphological characteristics of stem bending position, Crop Science Society of Japan, 4: 566-573 http://dx.doi.org/10.1626/jcs.60.566
Wang X., 2005, The thoughts of development quality super hybrid rice, Liaoning Nongye Kexue (Liaoning Agricultural Sciences), 3: 51-52
Wang Y., Li Q.Q., Li C.H., and Li A.F., 1998, Studies on the culm quality and anatomy of wheat varieties, Zuowu Xuebao (Acta Agronomica Sinica), 24(4): 452-458
Wang Y.R., Wang F.J., Yu J., Li Y., and Liu P.Q., 2010, Research progress on lodging resistance in rice, Guangxi Nongye Kexue (Guangxi Agricultural Sciences), 41(6): 545-547
Wolfgang S., Marc H.E., and Peter M., 2002, Semidwarf (sd-1), ‘‘green revolution’’ rice, contains a defective gibberellin 20-oxidase gene, Plant Biology, 99(13): 9043-9048
Xiao J.H., Li J.M., Yuan L.P., and Tanksley S.R., 1996, Identification of QTLs affecting traits of agronomic importance in a recombinant inbred population derived from a subspecific rice cross, Theor. Appl. Genet., 92(2): 230-244 http://dx.doi.org/10.1007/BF00223380
Xiao Y.H., Luo L.H., Yan X.Y., Gao Y.H., Wang C.M., Jiang L., Yano M., Zhai H.Q., and Wan J.M., 2005, Quantitative trait locus analysis of lodging index in rice (Oryza sativa L.), Zuowu xuebao (Acta Agronomica Sinica), 31(3): 348-354
Xing Z., Tan F., Hua P., Sun L., Xu G., and Zhang Q., 2002, Characterization of the main effects, epistatic effects and their environmental interactions of QTLs on the genetic basis of yield traits in rice, Theor. Aappl. Genet., 105(2-3): 248-257 http://dx.doi.org/10.1007/s00122-002-0952-y PMid:12582526
Yao F.X., ed., 2008, Experimental botany guide, Hubei Shifan Xueyuan Shengmingkexue Xueyuan, Hubei, China, pp.22-24
Ye S.P., Li J.Q., Zhang Q.J., Zhao B., and Li P., 2006, Mapping of quantitative traits loci for plant height of rice under different environments. Sichuan Nongye Daxue Xuebao (Journal of Sichuan Agricultural University), 24(1): 20-24
Zeng D.L., Hu J., Dong G.J., Liu J., and Qian Q., 2009, Quantitative trait loci mapping of flag-leaf ligule length in rice and alignment with zmlg1 gene, Journal of Integrative Plant Biology, 51(4): 360-366 http://dx.doi.org/10.1111/j.1744-7909.2008.00803.x PMid:21452587
Zhang F.Z., Jin Z.X., Ma G.H., Shang W.N., Liu H.Y., Xu M.L., and Liu Y., 2010, Dynamic changes of lodging resistance and chemical component contents in culm and sheaths of japonica rice during grain filling, Zhongguo Shuidao Kexue (Chin J RiceSci), 24(3): 264-270
Zhang H.C., and Wang F.Y., 2001, Recent progress on research of rice population in china, Zhongguo Shuidao Kexue (Chinese J Rice Sci), 15(1): 51-56
Zhang Q.Y., Ou-Yang Y.N., Dai W.M., Yu S.M., Zhuang J.Y., Jin Q.Y., and Cheng S.H., 2005, Relationship between traits of basal elongating internodes and lodging and QTL mapping in rice (Oryza sativa L.), Zuowu xuebao (Acta Agronomica Sinica), 31(6): 712-717
Zhang S.B., Zhao L., Zhu Z., Zhang Y.D., Chen T., Lin J., and Wang C.L., 2006, Genetic analysis for plant height of a semi-dwarf mutant 02428ha with eui gene in rice, Jiangsu Nongye Xuebao (Jiangsu J. of Agr. Sci.), 22 (2): 100-104
Zhuang J.Y., Fan Y.Y., Wu J.L., Xia Y.W., and Zheng K.L., 2001, Comparison of the detection of QTL for yield traits in different generations of a rice cross using two mapping approaches, Acta genetica Sinica, 28(5): 458-464



. PDF(299KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Huijuan Liu
. Yuchun Rao
. Yaolong Yang
. Yujia Leng
. Lichao Huang
. Guangheng Zhang
. Jiang Hu
. Longbiao Guo
. Zhenyu Gao
. Li Zhu
. Guojun Dong
. Jian Liu
. Meixian Yan
. Qian Qian
. Dali Zeng
Related articles
. Rice ( Oryza Sativa L spp. )
. Doubled haploid (DH)
. Quantitative trait loci (QTL)
. Leaf sheath
. Rice yield
Tools
. Email to a friend
. Post a comment